2024年3月7日,清华大学医学院、清华-IDG/麦戈文脑科学研究院、清华-北大生命科学联合中心郭增才受《神经元》杂志邀请在线合作发表题为《大脑左右半球交流:脑功能偏侧化之洞见》(Cross-hemispheric communication: Insights on lateralized brain functions)的综述论文,指出大脑功能的偏侧化不能完全用静态因素(如遗传差异、大脑结构的非对称性等)来解释,学习依赖的左右半球间的非对称信息交流对脑功能的偏侧化具有重要贡献。

从微小的淡水鱼到人类乃至更大的鲸类,所有脊椎动物的大脑都具有一个显著的特征,即大脑分为两个半球。左右半球并不相互孤立,它们通过多个白质纤维束相连,其中胼胝体是胎盘类哺乳动物的主要联络束。
在功能上,两个半球并不等效,几个重要的神经网络显示出功能的偏侧化。例如,在人类中,10.6%的人习惯用左手执行精细的运动如书写,而其余89.4%的人习惯用右手;这种精细运动的一侧偏好性在自然界普遍存在。除了运动领域,情绪处理也是脊椎动物脑功能偏侧化的一个重要领域。在人类中,对45种语言和12个语系进行的一项功能性磁共振成像研究发现,左侧化是人类语言网络的一个关键功能属性。神经解剖学上,语言生成和感知的Broca区和Wernicke区在超过95%人群中均在左半脑。
左右两个半球并不相互孤立,而是通过纤维束不断地交换信息。由于视觉、体感和听觉皮层区域优先处理来自对侧空间的信息,而运动皮层主要控制对侧身体的运动,跨半球通信在协调左右脑之间的感知、认知和运动功能中起着关键作用。在有些药物难以控制的癫痫病人中,病人不得已接受过手术操作以切断胼胝体(这些病人被称为裂脑人),术后发现裂脑人的很多认知功能具有偏侧化现象,通常左半脑在语言任务中起重要作用而右半脑在非语言和空间任务中有主导作用。
功能偏侧化是如何在神经生理层面上形成的?一个核心思想是跨半球交流导致功能偏侧化。从20世纪60年代对裂脑人的行为实验到现代神经影像学研究,许多关于左右脑信息交流的研究都是在人类参与者中进行的。然而,近年来随着在基因靶向、细胞类型特异性干扰、大规模电生理记录和定量行为评估方面的进展,啮齿类动物成为研究跨半脑通信极具吸引力的模型。重要的是,啮齿类动物模型可以在神经环路水平上提供深层机制的研究,可探索半脑间信息交流的因果性贡献,而这些通常在人类被试中难以实现。因此,本文旨在概述跨半脑信息交流,聚焦现代神经科技如何推动对跨半脑信息交流的理解,以及非对称信息交流如何推动脑功能偏侧化的诞生。
本文首先介绍左右半脑间信息交流的解剖基础,即纤维束(图1)。人类大脑中有三个较大的纤维束,包括前连合纤维束、胼胝体和海马连合纤维束。其中,前连合纤维束连接了颞叶皮层区域,参与嗅觉、疼痛知觉和生存;海马纤维束连接两个海马体,可能与记忆功能相关。胼胝体是连接皮层大部分区域的最大的纤维束,在人类中具有2-2.5亿根纤维,参与运动协调、半脑间感觉信息的整合、记忆痕迹的整合等。胼胝体的纤维通常连接同源区域,并具有大致的空间拓扑分布,即前部的纤维连接前部的大脑区域。根据连接的大脑区域,胼胝体又被分为几个不同的部分:前1/3区连接前额叶、前运动区和辅助运动区;前中部区连接运动区;后中部区连接体感和后顶叶区;峡部区连接上颞叶和后顶叶皮层;压部区连接下颞叶和枕叶皮层。这些组织形式在胎盘类哺乳动物里类似。值得注意的是,有袋类哺乳动物和鸟类中并没有胼胝体,它们的前连合纤维束在左右半脑皮层-皮层间的交流起更重要作用。

图1.人脑(上)和鼠脑(下)连接左右半脑的主要纤维束。前连合纤维束(右)、胼胝体(中)和海马连合纤维束(左)
接着该文介绍现代神经科学技术在研究左右半脑间信息交流中的应用。近年来,关于左右半脑信息交流的研究在啮齿类动物中大量涌现,这得益于现代神经科学技术在转基因动物中的使用。细胞类型特异的标记、大规模神经元活动记录、时空精确的干扰和定量行为分析等技术,能够用来研究左右半脑间的解剖连接、扰动左右半脑间的信息交流、或者直接记录左右半脑间信息交流相关的神经活动。本文介绍这些技术如何实现在单细胞水平上研究行为过程中左右半脑间的信息交流(图2)。
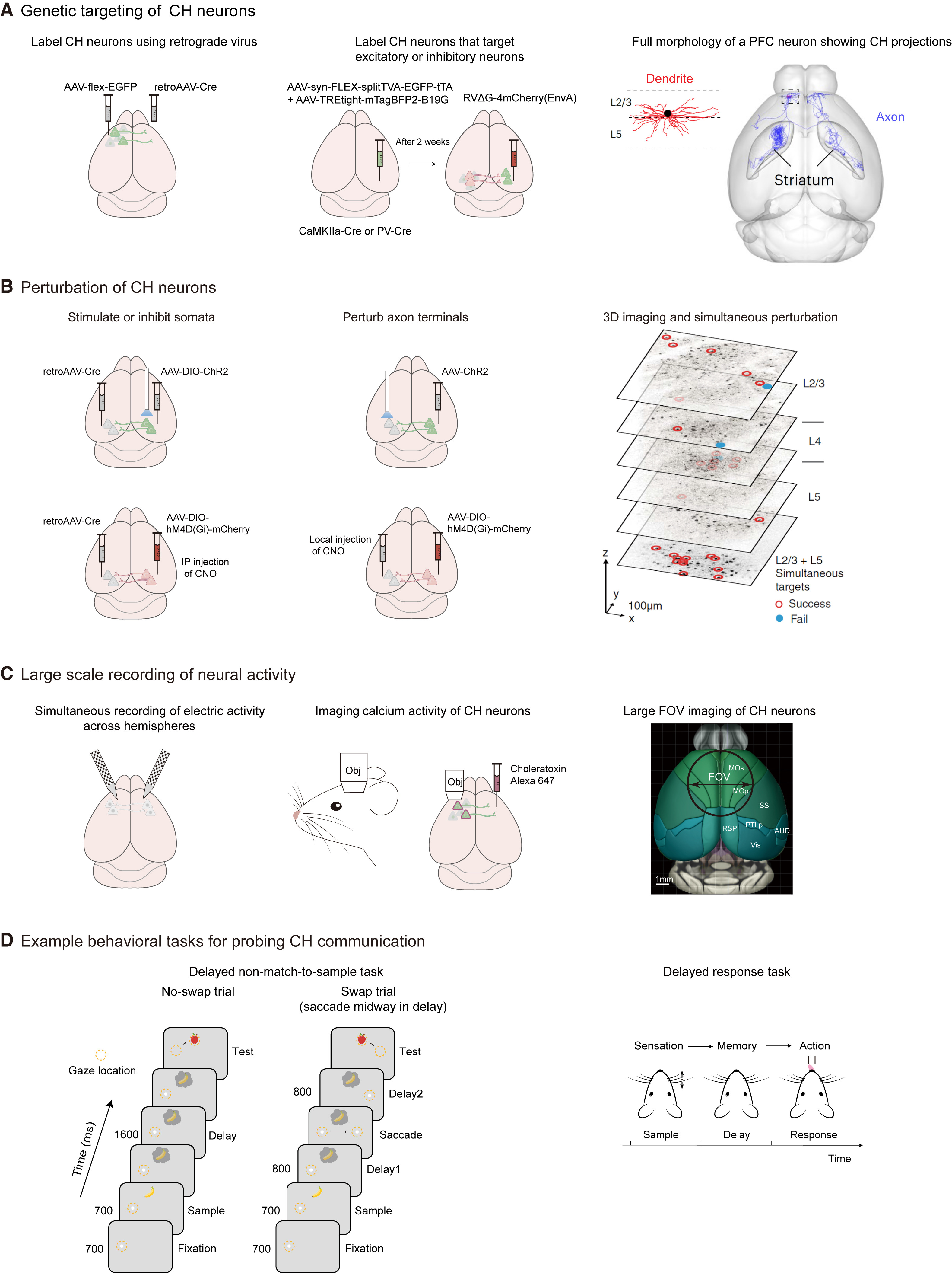
图2.采用现代神经科学技术细胞类型特异地标记半脑间交流神经元(CH neurons),从而在行为动物中时空精确地干扰或记录神经元群体活动。
(A)遗传标记半脑间神经元(左)或它们的半球间投射目标(中)。半脑间神经元通常与多个同侧和对侧脑区相连。右图显示了一个前额叶皮层第三层半球间投射神经元的完整形态。
(B)使用光遗传学和化学遗传学工具特异地干扰半球间投射神经元的策略(左)。通过局部给光或药物传递,干扰可以抑制半球间投射神经元的轴突末梢(中)。通过结合光遗传学和双光子成像技术,可以在三维成像过程中同时干扰群体神经元的活动(右)。
(C)大规模、多区域记录可以高通量地监测半球间投射神经元的活动(左,使用多个神经像素探针同时记录左右半脑的胞外电信号;中,使用双光子显微镜成像记录钙活动;右,大视场成像以覆盖具有半球间投射的脑区)。
(D)用于探索半球间信息交流的定量行为。左图为身体受限的猴子执行的非匹配视觉工作记忆任务。右图为头部固定的小鼠中的延迟反应任务。小鼠在采样阶段辨别触须刺激,在延迟阶段维持前瞻性短时记忆(运动计划),并根据感觉输入指示选择舔水口。
基于近期的研究,本文提出左右半脑间的信息交流表现为协调跨半球间的兴奋和抑制性输入。半脑间投射神经元大部分是兴奋性的,Ringo模型提出该投射提供兴奋性信息。由于较大的大脑信息传递的延迟变长,所以为降低该信息延迟的影响,越大的大脑应该有更强的功能偏侧化。尽管有部分结果与该模型相符,许多研究并没有发现大脑的尺寸和双手灵活性的关系。尽管半脑间投射是兴奋性的,在对侧半脑这些投射的对象通常包含许多抑制性神经元,理论上该通路可产生抑制性输入,很多电刺激实验也证实了这一点。但需要注意的是,这些扰动实验都是非自然地同时激活大量投射神经元。许多证据表明半脑间投射不仅传递兴奋性输入也传递抑制性输入,这表现在1)通常抑制一侧的皮层不会显著影响对侧皮层的整体神经活动,2)在决策行为中,抑制前运动皮层虽不影响对侧皮层的平均活动,但该操作特异地影响与行为相关的神经活动流形,这只有在协调兴奋性和抑制性影响的情况下才能实现。
左右半脑信息交流如何产生功能的偏侧化呢?发育学研究表明,固定的遗传倾向或脑结构的不对称通常只能解释个体偏好中的一小部分。这表明了动态过程对脑结构和功能不对称性起关键作用。例如,鸟类研究表明,在早期发育的关键时期,感觉环境对功能和结构侧化非常重要。具体而言,孵化期间的光照条件会改变鸟类的结构和功能不对称性,暗孵化会导致功能和结构不对称性的减少。光照孵化会影响即早基因的表达,并影响胚胎视网膜基因的表达。值得注意的是,最近在鸡的研究中也发现了基因与环境相互作用对半球间信息交流的影响。在该研究中,胚胎期的光刺激影响了半球间信息的传递:增强了从右半球到左半球的通信。这些发现突显了发展更加动态的观点来解释行为不对称性的重要性。
接下来,该文介绍了一个关于复杂相互作用的神经网络中半脑间信息交流的动态模型,指出半脑间不对称的信息交流导致处理感觉运动信息的功能偏侧化(图3)。感觉皮层倾向于优先处理来自对侧空间的信息,由于我们的注意力通常只能集中在一个主要物体上,将注意力引导到对侧半脑将增强对感觉刺激的检测。增强的感觉检测结合行为介导的奖励将进一步加强该侧半脑的感觉-运动关联。近年来基于触觉的决策任务的研究支持这一观点。小鼠胡须刺激可更快、更强地激活对侧前运动皮质,该区域呈现出对行为的主导性,而抑制主导的前运动皮层强烈地影响对侧非主导皮层的神经活动,反之,抑制非主导的半脑对主导半脑的决策活动影响较小。使用递归神经网络的模拟研究揭示,这种情况下半脑间的信息交流存在非对称性,信息从主导一侧的皮层流向非主导侧。因此,当强感觉刺激信息重复地传递到对侧半脑以指导运动时,学习依赖的可塑性可破坏对称的信息交流,导致大脑功能的偏侧化。
非对称的感觉刺激输入并不是诱导功能偏侧化的唯一因素。当同时刺激小鼠两侧的胡须时,前运动皮层仍可出现功能的偏侧化。在这种情况下,主导的一侧总是在强刺激关联的舔水方向的对侧,这表明感觉刺激和选择方向的关联决定了哪侧半脑主导。由于强刺激可以更容易被检测到,不显著的刺激引发的活动具有更低的信噪比,所以小鼠的最佳行为策略是优先强化强刺激关联的选择,而在弱刺激或无法检测到刺激时选择备份的方向。由于强刺激引发的动作高度依赖于对侧运动皮质,感觉-运动关联的加强将使对侧皮层占据优势。运动准备相关的活动在皮质中广泛分布,这种活动从主导的前运动皮层传递到对侧皮层,随着训练的进行,将导致对侧皮层对行为不在重要。因此,感觉-运动关联的不对称性也可以诱导半脑间非对称的信息交流和脑功能的偏侧化。最后,特定身体部位的重复动作可使对应的体感和运动皮层在体积上扩展。
理解半脑间信息交流和脑功能的偏侧化对相关领域也非常重要。例如,人脑中许多基本的认知功能都是非对称的,心理学研究可以从信息交流的非对称性中受益。此外,多种神经发育异常和精神障碍也与非正常的不对称性增加有关。近期的研究表明,机器学习算法和人工神经网络的编程受益于不对称性组织架构。因此,了解半脑间非对称信息交流的涌现与计算机科学的各个方面(包括人工智能的研究)相关。
清华大学医学院郭增才研究员、德国汉堡MSH医学院心理学系Sebastian Ocklenburg为本文通讯作者。该研究工作得到了科技部2030重大项目和国家自然科学面上基金的资助。郭增才课题组受到清华-北大生命科学联合中心和清华-IDG/麦戈文脑科学研究院的支持。
|
|